CONDITIONS FOR INFECTION OF WINTER OILSEED RAPE LEAVES BY CONIDIA AND ASCOSPORES OF PYRENOPEZIZA BRASSICAE, CAUSING LIGHT LEAF SPOT
Tijs Gilles1, Bruce DL Fitt1 and Michael J Jeger2
1IACR-Rothamsted, Harpenden, Herts, AL5 2JQ, UK
2Wageningen Agricultural University, Wageningen, 6709 PD, Netherlands
ABSTRACT
The optimum temperature for infection of oilseed rape leaves by P. brassicae conidia was c.16°C; an increase of temperature above the optimum and a decrease of temperature below the optimum decreased infection by conidia. Conidia did not infect oilseed rape leaves at 24°C. At the optimum temperature, conidia infected oilseed rape leaves after 6h of leaf wetness. At temperatures below the optimum, longer leaf wetness duration was required for infection by conidia. At 6°C, more than 16h of leaf wetness were required for conidia to infect oilseed rape leaves. The latent period of P. brassicae (time from inoculation to first sporulation) was shortest at c.16°C and increased as temperature increased above the optimum and below the optimum. Ascospores of P. brassicae were more infective than conidia on leaves of winter oilseed rape.
KEYWORDS: Brassica napus, Cylindrosporium concentricum, forecasts, infectivity, infection conditions, latent period
INTRODUCTION
Study of the conditions for infection of winter oilseed rape leaves by Pyrenopeziza brassicae (anamorph: Cylindrosporium concentricum) could contribute to the development of reliable weather-based forecast systems of light leaf spot on winter oilseed rape. Forecasts are needed to guide application of fungicides to control light leaf spot epidemics, which can cause severe yield losses (Fitt et al., 1997), because symptoms are often not apparent during autumn when fungicide sprays are most effective. If the relationships between environmental factors and infection are known, than the periods favourable for infection of winter oilseed rape crops could be estimated by the regular measurement of certain weather factors.
Previous studies have shown that temperature and leaf wetness duration influence infection of winter oilseed rape leaves by conidia. The length of the latent periods (time from inoculation to first sporulation) appeared to decrease linearly with increasing temperature from 5 to 15°C (Figueroa et al., 1995a). Less than 16h of leaf wetness inhibited infection of leaves by conidia at 12°C and 18°C (Figueroa et al., 1995b). However, the effects of temperature and leaf wetness duration on infection have never been studied over a wide temperature range.
P. brassicae produces two types of spores, namely conidia and ascospores. Conidia are asexually produced in acervular conidiomata, located in lesions on the plant surface (Rawlinson et al., 1978); ascospores are produced during the sexual stage in apothecia on debris (Lacey et al., 1987; McCartney and Lacey, 1990). Although conidia and ascospores are morphologically identical when seen with a light microscope (Rawlinson et al., 1978), their infectivities and thus their importance in the development of light leaf spot epidemics may differ.
This paper reports new information about the conditions for infection of winter oilseed rape leaves by conidia, compares the infectivity of conidia and ascospores, and discusses how this knowledge of conditions for infection could be used to develop weather-based forecast systems of light leaf spot on winter oilseed rape.
MATERIAL AND METHODS
Conditions for infection of oilseed rape leaves by conidia
Winter oilseed rape plants (cv. Bristol) were grown at 16°C for 3.5 weeks until 4-5 expanded leaves had been developed. Two days before inoculation, plants were transferred to controlled environment cabinets at 4, 6, 8, 12, 16, 20 or 24°C to acclimatise. The plants were inoculated with a suspension of 106 conidia per ml of a field isolate of P. brassicae, derived from sporulating infected leaves at Rothamsted in May 1997. Directly after inoculation, plants were enclosed in polyethylene bags to maintain leaf wetness. The bags were removed after 6, 10, 16, 24, 48 or 72 h, and leaves had dried within 30 minutes. % leaf area with sporulation was assessed visually on leaves 3 and 4 (oldest leaf is leaf 1) of five plants for each leaf wetness duration and temperature treatment combination. The infection experiment was replicated three times over a period of 6 months.
Comparison of infectivity of conidia and ascospores
Winter oilseed rape plants (cv. Bristol) at the 5-leaf-stage (3.5 weeks old) were inoculated with either conidia or ascospores of P. brassicae at different densities of spores per unit leaf area, under the same conditions during infection (16°C, 24 h leaf wetness). The plants were inoculated with conidia by spraying conidial suspensions with an aerosol-powered sprayer onto the plants for equal lengths of time. By spraying suspensions of different conidial concentration onto the plants, conidia were deposited at different densities per unit leaf area. Suspensions of 106, 105, 104, 103 or 102 conidia ml-1 were sprayed onto the plants. Plants were inoculated with ascospores from mature apothecia located above the plants. Exposing the plants to ascospore release for 0.5, 1, 1.5, 2 or 3h deposited ascospores onto the plants at different densities of ascospores per unit area of leaf.
Infectivity was calculated as the number of lesions produced per number of spores deposited onto a leaf. During inoculation, microscope slides had been placed around the plants to assess the number of spores deposited per unit area. The number of spores per unit area of slide multiplied by the units area of leaves at the time of inoculation gave an estimate of the number of spores deposited onto a leaf. Two weeks after inoculation, numbers of lesions were assessed on leaves 3 and 4 of each plant.
RESULTS
Conditions for infection of oilseed rape leaves by conidia
Temperature was optimal for infection of winter oilseed rape (cv. Bristol) by conidia of P. brassicae at c.16°C (Figure 1). At 16°C and 48h leaf wetness duration, the latent period was shortest (10 days) and the greatest % area of leaves produced spore pustules (42%). An increase of temperature above the optimum and a decrease of temperature below the optimum decreased % leaf area with sporulation. No sporulation was observed at 24°C. Leaf wetness duration interacted with temperature. Increasingly longer leaf wetness was needed for infection at temperatures decreasing below the optimum. At 16°C, little infection occurred after 6 h of leaf wetness, but at 6°C more than 16 h of leaf wetness was required for conidia to infect the leaves.
The latent period was affected by temperature (Figure 2) and was shortest at 16°C. When temperature increased above or decreased below the optimum, the latent period increased.

6 h
Figure 1. Effects
of temperature and leaf wetness duration on % leaf area with sporulation after
inoculation of winter oilseed rape (cv. Bristol) with conidia (average of 3 replicates).
|
6 h |
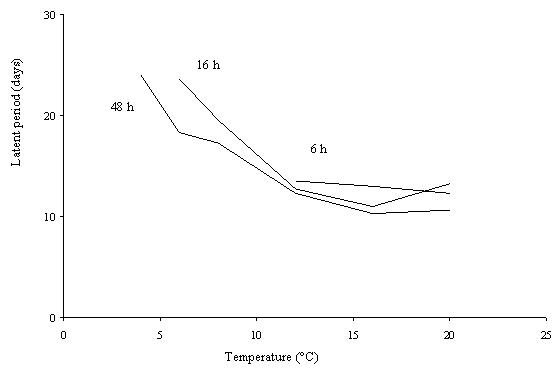 |
Figure 2. Latent periods (time from inoculation to first sporulation) observed after inoculation of leaves of winter oilseed rape (cv. Bristol) with conidia at different temperatures and leaf wetness duration (average of 3 replicates).
Comparison of infectivity of conidia and ascospores
The infectivity of ascospores of P. brassicae differed from the infectivity of conidia of P. brassicae. In comparison, on average 100 lesions per leaf were caused by 4000 to 5000 ascospores per leaf or by approximately 2000000 conidia per leaf. In most cases no lesions were caused by less than approximately 50000 conidia per leaf. Ascospores, however, caused lesions when less than 500 ascospores had deposited on a leaf.
DISCUSSION
The knowledge of conditions for infection could used to estimate the periods when conditions are favourable for primary infections by ascospores, secondary infections by conidia and late-secondary infection by ascospores by the regular measurement of weather data. Ascospores are likely to cause primary infections in the autumn in winter oilseed rape crops more often than conidia, and could contribute to stem and pod infections in spring and summer. Ascospores are likely to be frequently produced in the UK (Ball et al., 1990; Majer et al., 1998). Because of their wind-dispersal (McCartney et al., 1986), ascospores are more likely to be dispersed from a distant inoculum source and deposited in a winter oilseed rape crop as primary inoculum in the autumn than the splash-dispersed conidia (Rawlinson et al., 1978). Furthermore, their greater infectivity implies that once deposited onto leaves in a crop, ascospores are more likely to cause primary infection than conidia. Ascospores released from leaf debris in crops in spring and early summer (McCartney and Lacey, 1990) could cause frequently observed stem and pod infections. Conidia are likely to cause secondary infections after primary infection in the autumn at short distances from their source, the sporulating lesions. The dispersal by rain-splash of conidia enables them to be dispersed over short distances from plant to plant and within plants (Rawlinson et al., 1978).
Current knowledge of the conditions for infection by conidia can be modelled into a function describing the relation leaf wetness duration and temperature and infection, but more information is needed of the conditions for infection by ascospores. The relation between latent period and temperature could be approximated with a linear curve for the temperature range 5-15°C as mentioned by Figueroa et al. (1995a). However, a non-linear curve seems more appropriate for the temperature range 4-20°C. Infection was inhibited at 24°C. Thus, at warm days in autumn, spring and summer infection of leaves by conidia is inhibited. Because infection was not inhibited at 4°C, effects of even lower temperatures on infection by conidia need to be investigated. Figueroa et al. (1995b) found that a duration of leaf wetness shorter than 16h inhibited infection. Results of present study suggest that at temperatures around the optimum of 16°C, conidia can infect winter oilseed rape leaves after less than 16h of leaf wetness; 6h of leaf wetness was found to be sufficient for conidia to infect at 16°C.
ACKNOWLEDGEMENTS
We acknowledge the Perry Foundation and the UK Ministry of Agriculture Fisheries and Food for their financial support.
REFERENCES
Ball AM, Siddiq AA and Giltrap NJ (1990) Assessment of benomyl resistance and mating type in field isolates of Pyrenopeziza brassicae, cause of light leaf spot of brassicas. Plant Pathology 39, 33-37.
Figueroa L, Fitt BDL, Shaw MW, McCartney HA and Welham SJ (1995a) Effects of temperature on the development of light leaf spot (Pyrenopeziza brassicae) on oilseed rape (Brassica napus). Plant Pathology 44, 51-62.
Figueroa L, Fitt BDL, Welham SJ, Shaw MW and McCartney HA (1995b) Early development of light leaf spot (Pyrenopeziza brassicae) on winter oilseed rape (Brassica napus) in relation to temperature and leaf wetness. Plant Pathology 44, 641-654.
Fitt BDL, Gladders P, Turner JA, Sutherland KG, Welham SJ and Li Davies JM (1997) Prospects for developing a forecasting scheme to optimise use of fungicides for disease control on winter oilseed rape in the UK. Aspects of Applied Biology 48, 135-142.
Lacey ME, Rawlinson CJ and McCartney HA (1987) First record of the natural occurrence in England of the teleomorph of Pyrenopeziza brassicae on oilseed rape. Transactions of the British Mycological Society 89, 135-140.
Majer D, Lewis BG and Mithen R (1998) Genetic variation among field isolates of Pyrenopeziza brassicae. Plant Pathology 47, 22-28.
McCartney HA and Lacey ME (1990) The production and release of ascospores of Pyrenopeziza brassicae on oilseed rape. Plant Pathology 39, 17-32.
McCartney HA, Lacey ME and Rawlinson CJ (1986) Dispersal of Pyrenopeziza brassicae spores from an oil-seed rape crop. Journal of Agricultural Science, Cambridge 107, 299-305.
Rawlinson CJ, Sutton BC and Muthyalu G (1978) Taxonomy and biology of Pyrenopeziza brassicae sp.nov. (Cylindrosporium concentricum), a pathogen of winter oilseed rape (Brassica napus ssp. oleifera). Transactions of the British Mycological Society 71, 425-439.