INTEGRATED MANAGEMENT OF ROOT MAGGOTS (DIPTERA)
IN CANOLA
Lloyd M. Dosdall
Alberta Agriculture, Food and Rural Development, 7000-113 Street, Edmonton, Alberta Canada T6H 5T6; e-mail: lloyd.dosdall@agric.gov.ab.ca
ABSTRACT
In central Alberta, Canada, infestations of the root maggots, Delia radicum (L.) and Delia floralis (Fallén), are responsible for substantial economic losses in the production of canola. Primary plant damage is caused by larvae feeding on root tissue, and secondary damage occurs when feeding channels are subsequently invaded by root rot fungi. This may result in host plant mortality, or decreased crop yields estimated in central Alberta at 52% for Brassica rapa L. and 20% for Brassica napus L. Recent research toward development of an integrated management strategy for root maggots in canola has focused on cultural strategies, insecticidal control, and host plant resistance. To reduce economic losses from root maggot attack, canola growers should plant B. napus rather than B. rapa in regions where the growing season is sufficiently long, and they should also select the least susceptible cultivars of those species. Cultivation prior to seeding reduces the overwintering survival of root maggot puparia. Planting canola at seeding rates higher than those currently recommended, and sowing seed with organophosphate granules coated with a synthetic polymer for slow release in the soil reduces crop damage and can result in improved seed yields. Research to develop canola germplasm with resistance to root maggots has identified sources of resistance, and plant biotechnological approaches are being used to incorporate these genes in commercial cultivars of canola.
KEYWORDS: Delia spp., cultural control, tillage, insecticides
INTRODUCTION
In central Alberta, Canada, infestations of the root maggots, Delia radicum (L.) and Delia floralis (Fallén) (Diptera: Anthomyiidae), and secondary plant damage by root rot fungi (Fusarium spp.), can cause decreased crop yields estimated at 52% for Brassica rapa L. and 20% for Brassica napus L. (Griffiths 1986, 1991). Although the literature contains several reports of effective chemical, cultural, and biological control strategies for reducing the impact of root maggot infestations in market garden cruciferous crops, these strategies are not appropriate for canola in western Canada for several reasons. Treatment of adults with insecticidal sprays is impractical because emergence occurs over an extended period, and larvicides are not feasible because of poor spray penetration of the crop canopy in mid-summer (Griffiths 1986). Moreover, insecticides adversely affect predators and parasites of root maggots (Coaker and Williams 1963), and their repeated use has caused resistance in some populations of D. radicum (Coaker et al. 1963). Cultural practices, such as the use of row covers as described by Matthews-Gehringer and Hough-Goldstein (1988), are excessively labor-intensive given the vast areas seeded to the crop and the relatively low economic value of each canola plant. Several pathogens, predators, and parasites may reduce root maggot populations, but little potential exists for biological control agents acting alone to substantially reduce economic damage in canola. Consequently, the research described here has focused on developing an integrated management strategy for root maggots in canola that combines effective and practical cultural and chemical strategies. The susceptibilities of different canola species and cultivars of each species were determined with a view to identifying those least susceptible to infestation, and different tillage regimes were studied for reducing emergence of root maggot adults. Seeding rate and seeding date were assessed in conjunction with applications of organophosphate insecticides, coated for slow release in the soil and applied with the seed, for the integrated management of Delia spp. Populations in canola.
METHODS AND MATERIALS
Root maggot infestations were investigated on three cultivars of B. rapa (cvs. Horizon, Colt, and Parkland), three cultivars of B. napus (cvs. Legend, Bounty, and Celebra), seven cultivars of Brassica juncea (cvs. DN 1738, Donskaja, Common Brown, Scimitar, Lethbridge 22A, Domo, and Cutlass), and two cultivars of Sinapis alba (cvs. Ochre and Tilney). The experiments were randomized complete-block designs with three replications. Experimental plots measured 6 by 1 m and comprised four rows spaced 20 cm apart. Susceptibilities of species and cultivars to infestation by Delia spp. were assessed by evaluating root injury at the end of the growing season using the semi-quantitative rating scale of Dosdall et al. (1994). Significance of differences in root damage ratings among and within species were determined using analysis of variance (P £ 0.05) and Student-Newman-Keuls multiple comparisons (SAS Institute Inc. 1990), after performing log10 (x + 1) transformations on the data.
The effect of tillage regime on emergence of root maggot adults was determined using a randomized split-plot design with canola species (B. rapa cv. Tobin and B. napus cv. Alto) assigned to main plots and tillage treatment assigned to sub-plots. There were four replications and sub-plots measured 15 by 15 m. Tillage treatments evaluated were: tillage in the fall only, tillage in the spring only, tillage in both the fall and the spring, and no tillage. Tillage involved cultivating with tandem discs and mounted tine harrows so the soil was worked to a depth of approximately 15 cm. In spring, four emergence traps were placed randomly on each sub-plot. Each emergence trap sampled an area of 1 m2. Twice weekly from May to July, all newly eclosed insects were collected from the emergence traps. Data were subjected to analysis of variance (P £ 0.05) and Duncan's New Multiple Range Test (SAS Institute Inc. 1990) to determine the significance of differences in emergence of root maggot adults for the various tillage regimes.
Experiments to assess integrated effects of seeding date, plant density, and insecticidal seed treatment were randomized split-plot designs with four replications conducted at two sites in each of two years. Seeding dates (early May, mid May, and late May) were assigned to main plots and plant density (35.6 ± 1.6 [S.E.], 40.9 ± 1.8 [S.E.], and 45.0 ± 1.7 [S.E.] plants per m row), and chemical seed treatment (isofenphos or Amaze® at 12 mL per kg, terbufos or Counter® in a 1:1 mixture with the seed, and carbathiin:thiram:lindane or Vitavax rs®) were assigned to sub-plots. Seed treatments with isofenphos and terbufos included an outer coating of a synthetic polymer to delay release of the insecticide into the soil. Sub-plots measured 6 by 1 m, and comprised four rows spaced 20 cm apart. Root maggot infestations were assessed by root damage at the end of the season according to the method described previously. Means were compared using analysis of variance (P £ 0.05) and Student-Newman-Keuls multiple comparisons (SAS Institue Inc. 1990), after performing log10 (x + 1) transformation on the data.
RESULTS
Differences in susceptibility to infestation by Delia spp. occurred among and within canola and mustard species. In terms of root damage, the order of susceptibility of these crops was B. rapa > B. juncea > B. napus > S. alba. The most susceptible cultivars of B. rapa were Horizon and Colt; of the B. napus cultivars studied, all had similar susceptibilities to root maggot attack. Brassica juncea cv. DN 1738 was susceptible and more similar in susceptibility to B. rapa cultivars than to other cultivars of B. juncea. Susceptibility was similar for the two S. alba cultivars studied.
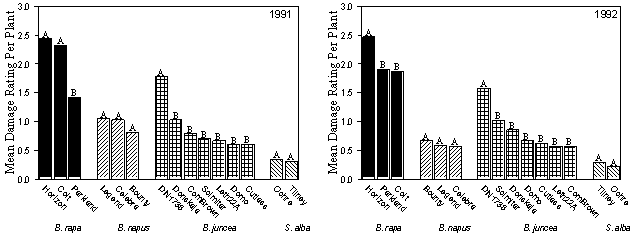 |
Fig. 1. Mean damage ratings per plant for various cultivars of Brassica rapa, B. napus, B. juncea, and Sinapis alba. Letters on histograms indicate significance of differences between cultivars within each species; means within a species having the same letter indicate no significant differences using ANOVA and Student-Newman-Keuls multiple comparisons.
Tillage effects on emergence were highly significant for both years of the study and for both D. radicum and D. floralis (P < 0.01). Emergence of D. radicum was significantly greater from plots that had not been tilled relative to plots subjected to any of the other tillage treatments (P < 0.05), but no significant differences in emergence were observed among plots tilled only in the spring, only in the fall, and in both the fall and the spring. Emergence of D. floralis was also significantly greater from untilled plots compared with those subjected to any other tillage treatment (P < 0.05), but significantly more D. floralis emerged from plots tilled only in the spring than from plots tilled only in the fall and in both the fall and the spring.
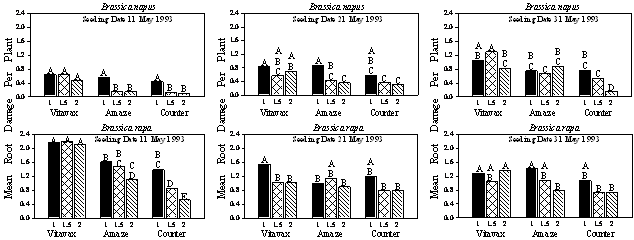 |
Fig. 2. Mean damage ratings per plant for B. napus and B. rapa seeded on different dates, at different rates (1, 1.5, and 2 times the recommended rate), and with various seed treatments (Vitavax, Amaze, and Counter). Letters on histograms indicate significance of difference between treatments: means within a species and seeding date having the same letter indicate no significant differences using ANOVA and Student-Newman-Keuls multiple comparisons.
Studies of integrated effects of seeding date, seeding rate, and insecticidal seed dressing determined that root damage was greater on B. rapa plants than on B. napus for all seeding dates (Fig. 2). For B. rapa, root damage was greater for plants treated with Vitavax rs® and seeded on the earliest date than for plants subjected to other treatments. For plants seeded later in the season, root damage tended to be greater at the lowest plant density. Lowest root damage was recorded for plants treated with isofenphos (Amaze®) and terbufos (Counter®) at the highest plant density. Brassica napus plants seeded on the latest date generally had greater root injury than those seeded earlier in the season. However, for practically all treatments, root damage declined with an increase in plant density. Lowest root damage values were observed for plants treated with terbufos (Counter®) and grown at the highest density.
DISCUSSION
Because of the lower susceptibility of B. napus than B. rapa to infestation by root maggots, canola producers should seed B. napus in areas infested annually by high population densities of Delia spp., and where the growing season is of sufficient length (110-120 days) for crops to reach maturity. Where the season is shorter (90-100 days), less susceptible cultivars of B. rapa (e.g. cv. Parkland) should be selected rather than more susceptible ones (e.g. cv. Horizon). The comparatively greater resistance of S. alba than the other canola and mustard species evaluated here indicates its potential as a genetic source of resistance to root maggots. Research is now in progress to use biotechnological methods to identify and transfer genes for root maggot resistance from S. alba to commercial cultivars of canola.
Regardless of the timing of its implementation, tillage was effective for reducing root maggot emergence from plots seeded previously to B. rapa and B. napus. There are several possible mechanisms by which tillage could reduce emergence of Delia spp. In addition to causing mechanical injury to puparia, tillage can move puparia nearer the soil surface where they would be exposed to more severe environmental conditions, especially during winter. Near the soil surface they would also be more exposed to attack by predators and parasites (Stinner and House 1990).
When the integrated effects of seeding date, plant density, and chemical seed treatment were evaluated for each canola species, it was apparent that canola species was the single most important factor in limiting infestations by Delia spp. For practically any given seeding date, plant density, and chemical seed treatment, damage to plants of B. rapa exceeded that to B. napus. Plant density and chemical seed treatment were also important for reducing infestations by Delia spp. Root damage was generally greatest on plants seeded at lower densities than at higher densities. Plants subjected to seed treatments with either isofenphos or terbufos that were coated with a synthetic polymer for slow release in the soil generally had less root damage than plants which developed from seed treated with Vitavax rs®. Use of a terbufos seed treatment applied to plants grown at a high density appears to be the most effective control practice for Delia spp. in canola.
Although early sown crops benefit from a full season's rainfall, suffer less weed competition, and benefit from the higher soil nitrate levels available at the beginning of the season (Dent 1991), results of this study indicated that delaying seeding date until late May (rather than early or mid-May) resulted in less root injury to both B. rapa and B. napus. However, delayed seeding was also associated with substantial reductions in seed yield; evidently, early-seeded crops could better compensate for increased root maggot damage. Delaying seeding date is therefore not an appropriate cultural control practice for root maggots.
The impact of root maggot infestations in canola in western Canada can be reduced through implementation of some strategies investigated in these studies. Seeding species and cultivars least susceptible to infestation can reduce larval damage to taproots. Cultivation prior to seeding reduces emergence success of root maggot adults. Seeding at rates higher than those recommended in the absence of root maggot attack, and application of a seed treatment coated with a synthetic polymer coated for slow release in the soil can also limit the impact of root maggot infestations. The most environmentally sustainable approach for root maggot control would be to develop crop cultivars resistant to root maggot attack, and research is currently focused in this direction.
ACKNOWLEDGEMENTS
This research was funded by the Alberta Canola Producers Commission, the Alberta Agricultural Research Institute, and the Alberta Research Council. For technical assistance, I am very grateful to N.T. Cowle, P.M. Conway, M.J. Herbut, and T.M. Micklich.
REFERENCES
Coaker, T.H., and D.A. Williams. 1963. The importance of some Carabidae and Staphylinidae as predators of the cabbage root fly, Erioischia brassicae (B.). Entomologica Experimentalis et Applicata 6: 156-164.
Coaker, T.H., D.J. Mowat, and G.A. Wheatley. 1963. Insecticide resistance in the cabbage rootfly in Britain. Nature 200: 664-665.
Dent, D. 1991. Insect Pest Management. C×A×B International, Wallingford, Oxon, U.K. 604 pp.
Dosdall, L.M., M.J. Herbut, and N.T. Cowle. 1994. Susceptibilities of species and cultivars of canola and mustard to infestation by root maggots (Delia spp.) (Diptera: Anthomyiidae). The Canadian Entomologist 126: 251-260.
Griffiths, G.C.D. 1986. Phenology and dispersion of Delia radicum (L.) (Diptera: Anthomyiidae) in canola fields at Morinville, Alberta. Quaestiones Entomologicae 22: 29-50.
Griffiths, G.C.D. 1991. Economic assessment of cabbage maggot damage in canola in Alberta. Proceedings of the GCIRC Eighth International Rapeseed Congress (Saskatoon, July 9-11, 1991) 2: 528-535.
Matthews-Gehringer, D., and J. Hough-Goldstein. 1988. Physical barriers and cultural practices in cabbage maggot (Diptera: Anthomyiidae) management on broccoli and Chinese cabbage. Journal of Economic Entomology 81: 354-360.
SAS Institute Inc. 1990. SAS System for Personal Computers. Release 6.04. SAS Institute Inc., Cary, NC.
Stinner, B.R., and G.J. House. 1990. Arthropods and other invertebrates in conservation-tillage agriculture. Annual Review of Entomology 35: 299-318.