CHARACTERISTICS OF DOUBLE LOW WINTER RAPESEED LINES WITH INTRODUCED RESTORER GENE FOR CMS OGURA
Iwona Bartkowiak-Broda, Wiesława Popławska
Plant Breeding and Acclimatization Institute (IHAR) - Strzeszyńska 36,
60-479 Poznań, Poland, E-mail: ibart @ nico.ihar.poznan.pl
ABSTRACT
The main purpose of the investigations was to obtain the restorer lines for CMS ogura system with low glucosinolate content and good yielding ability.
The plants used for the study were selected in F3 generation of hybrids between CMS lines with low glucosinolate content or double low fertile lines and restorer lines with glucosinolate content of about 60 µM/g of seeds. The 30 selected plants homozygous in respect of restorer gene alleles were characterized by content of total glucosinolates in the range of 10,7 - 19,6 µM/g of seeds. The yield components (number of seeds per pod, 1000 seed weight) revealed high variability.
The obtained results showed that in genome of the restorer lines with low glucosinolate content the isozyme marker PGI - 2 and PCR-RAPD molecular marker OPC 02 are not lost with the fragment of DNA responsible for high glucosinolate content.
KEYWORDS: glucosinolates, yield components, molecular markers
INTRODUCTION
The development of hybrid varieties of rapeseed with the use of CMS ogura has been one of the main breeding objectives in the past few years. The lack of restorer lines with appropriate qualitative and agronomical traits is the factor limiting the utilization of CMS ogura in breeding of restored hybrid varieties. This difficulty is due to the origin of restorer lines in Brassica napus. Restorer gene has been introduced to the rapeseed genome from radish (Raphanus sativus) genotype (Heyn 1976). Obtained recombinants were characterized by low seed set in pods attributed to a high rate of embryo sac abortion (Pellan - Delourme and Renard 1988; Delourme et al 1991) and strong linkage of restorer alleles with genes determining high glucosinolate content (Delourme et al 1995). These recombinants retained too much genetic information from radish. Investigations conducted by Delourme et al (1995) revealed that the improvement of these traits is possible by backcrosses with double low lines and elimination of radish genetic information.
The aim of the investigations undertaken by Oil Crop Department of IHAR was to obtain restorer lines with low glucosinolate content and good yielding ability.
MATERIAL AND METHODS
Material for investigations was constituted of:
- restorer line R obtained from INRA - France within the licence agreement, heterozygous in respect of restorer gene alleles, with total glucosinolate content of about 60 µM/g of seeds,
- double low lines of winter rapeseed with glucosinolate content 5,1 - 11,8 µM/g of seeds,
- CMS ogura lines with glucosinolate content 12,2 - 16,0 µM/g of seeds.
Investigations were carried out in F3 generation selected from hybrids between restorer line R and low glucosinolate lines with sterile or normal cytoplasm.
Selection of genotypes with restorer gene alleles was carried out on the phenotypic expression and with the use of isozyme marker PGI-2 (Delourme and Eber 1992). The presence of molecular marker of restorer gene RAPD-OPC02 (Delourme et al 1994) was investigated only in lines characterized by low glucosinolate content. The analyses of glucosinolates were performed with the method of gas chromatography of silyl derivatives of desulfoglucosinolates. Seed set in pods and 1000 seed weight were evaluated in F3 progeny on ten plants from each of the 44 examined lines.
RESULTS
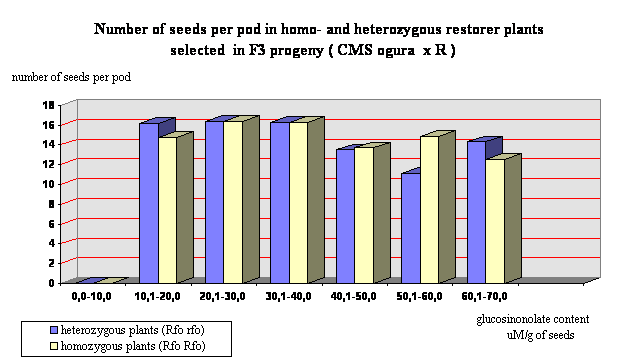
Segregating population F2 of about 2500 plants were obtained by crossings of starting restorer line with CMS ogura and double low lines with normal cytoplasm. From this population 44 lines were selected. They were characterized by glucosinolate content from 13,6 µM/g of seeds to 25,6 µM/g of seeds and relatively regular meiotic behaviour, but still disturbed by the presence of some univalents and in some cases multivatents. In F3 progeny obtained from these lines 429 male fertile plants were selected for futher investigations. After examination with the use of PGI-2 marker it was observed that 215 plants were homozygous (Rfo Rfo) and 214 plants were heterozygous (Rfo rfo). The coefficient of variability in respect of glucosinolate content was very high for both groups of plants (Figure 1), but 30 homozygous plants with total glucosinolate content not exceeding 20 µM/g of seeds were selected. The main yield component number of seeds per pod was not dependent on glucosinolate content (Figure 2). The correlation coefficient between these two traits was statistically not significant (r = - 0,117). However, the mean value of seed set per pod for homozygous plants was 15,88 and for heterozygous plants 15,62. It was below the mean value for standard variety which amounted in the same growing conditions 22,4, but several plants not exceeding 15 µM/g of seeds of glucosinolates (Polish norm for sowing material) revealed seed set per pod in range 15 to 24,5.
The presence of PGI-2 and RAPD OPC02 markers was observed in all low glucosinolate homo- and heterozygous genotypes.
CONCLUSION
The selected double low restorer lines with improved productivity will be used to produce restored F1 hybrids.
Two investigated markers PGI-2 and OPC02 closely linked to the restorer gene are not lost in low glucosinolate recombinants together with the fragment of DNA responsible for high glucosinolate content.
Figure 1
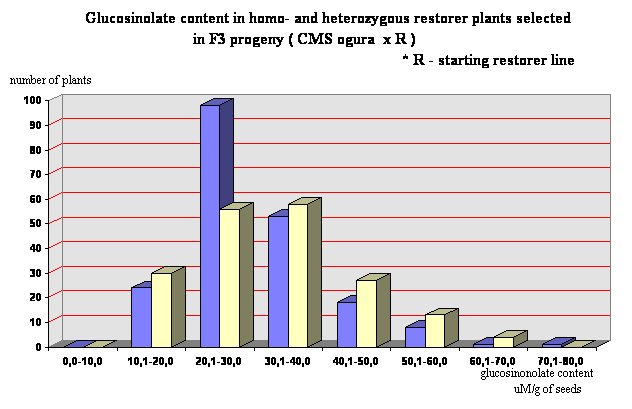
Figure 2